Plantepatogene sopp har evnen til å utvikle resistens eller redusert følsomhet overfor fungicider/soppmidler som brukes for å bekjempe dem. Fungicider slutter å gi tilstrekkelig kontroll av sykdommer forårsaket av sopp hvis resistens bygges opp i populasjonen. Soppens biologi, fungicidets virkemåte og agronomiske faktorer påvirker risiko for utvikling av fungicidresistens.
Navigating the threat of azole resistance development in human plant and animal pathogens (NavAzole) 2021-2025 (NFR nummer: 320821)
RESISTOPP: Plantevernmiddelresistens: Mutasjon, seleksjon og spredning (NIBIO SiS), 2017-2021, prosjektleder: Arne Stensvand
Gråskimmel på bringebærstengel. Foto: Jafar Razzaghian
Fungicidresistens er arvelig
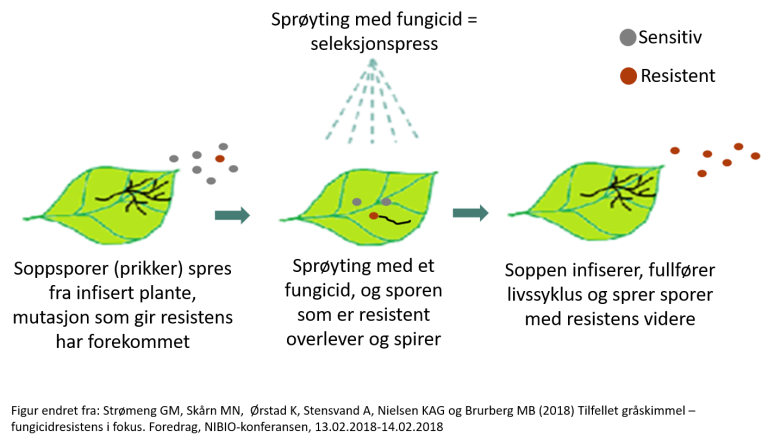
Fungicidresistens er en arvelig reduksjon i følsomhet av en sopp overfor et soppbekjempende middel (fungicid). Mange populasjoner av patogene sopp har naturlig variasjon i følsomheten mot fungicider mellom individer.
Dette betyr at før et fungicid er tatt i bruk er det mulig at det eksisterer individer som tåler fungicidet eller er mindre følsom enn andre. Spontane endringer i arvestoffet, mutasjoner, kan oppstå hos individer i populasjoner også. I noen tilfeller kan slike spontane mutasjoner gjør at et individ tåler et fungicid i større grad enn andre i samme populasjonen.
For hver gang et fungicid blir brukt, skjer det en seleksjonsprosess der mottakelige individer blir drept eller hemmet av fungicidet og individer som tåler fungicidet overlever og er selektert. Resistente individer som overlever fungicidbehandling, er også de som kan oppformere seg videre når forholdene tillater det.
Fungicidresistens er arvelig, så overlevende individer vil produsere resistente avkom. Videre bruk av fungicidet vil gjøre at andelen resistente individer i en populasjon vil fortsette å øke til fungicidet har mistet effektiviteten og ikke lenger kan brukes til å kontrollere sykdommen tilstrekkelig.
Hvordan måles resistens eller redusert følsomhet?
Måling av resistens eller redusert følsomhet begynner med prøveuttak og isolering av sopp fra infisert plantemateriale. En måte å teste soppisolater for følsomhet mot fungicider på er å overføre soppisolater til vekstmedier tilsatt ulike konsentrasjoner av fungicidet (fortynningsserie), og måle relativ vekst i forhold til vekst på medier uten fungicidet.
Resultatene brukes for å regne ut en EC50-verdi, som er den konsentrasjonen av fungicidet som hemmer veksten 50 prosent i forhold til vekst på medier uten tilsatt fungicid. Ved å sammenligne EC50-verdier for soppisolatene som testes med EC50-verdier fra en sensitiv populasjon som ikke har vært utsatt for fungicidet, kan man se om det har blitt endring i følsomhet hos soppen. Det er avgjørende å inkludere referanseisolater med kjente resistensprofiler for å kunne sammenligne resultater fra ulike tester.
EC50-verdier brukes for å regne ut en resistens-faktor (RF) ved å dele EC50-verdien av et isolat som testes med EC50-verdien til det følsomme referanseisolatet.
RF-verdier viser hvor mye følsomheten hos soppen har endret seg i forhold til en standard.
Avhengig av hvilke resistensmekanismer og hvor mange gener som er involvert, kan endring i EC50-verdier skjer gradvis over tid, eller vise raskt og drastisk økning.
I noen tilfeller, kan data om EC50-verdier også brukes for å velge diskriminerende doser av fungicider. En diskriminerende dose er en fungicidkonsentrasjon som kan benyttes for å skille mellom resistente og sensitive individer i en resistenstest.
Testing med diskriminerendedoser er en raskt og enkel metode for å få oversikt over resistenssituasjonen hos en populasjon av sopp i et område. Det er mer effektivt å teste kun én konsentrasjon av et fungicid fremfor å bruke en fortynningsserie.
Likevel kan en diskriminerende dose ikke fange opp små endringer i EC50-verdier hos soppen. Bruk av diskriminerende doser kan egne seg til overvåkningsarbeid der raskt resultat er viktig mens bruk av fortynningserier og beregning av EC50-verdier vil gi mer detaljerte informasjon om endringer i følsomheten hos soppen over tid.
Betydning av endringer i EC50-verdier hos soppen for effektiviteten av fungicidet mot soppen ute i felt er viktig å undersøke. En test der effekten av fungicidet testes på sopp på plantemateriale (bioassay) gir den beste indikasjonen på om endring i følsomhet har praktisk betydning for kontroll av soppsykdommen ute i felt. Bioassay-metoden er også den mest ressurs- og tidskrevende måte å teste på.
Bruk av molekylære metoder for å påvise mutasjoner som gir resistens hos soppen har blitt mer aktuelt i de siste årene. Hurtige molekylære metoder kan være nyttige for overvåkning av sopparter der resistensmekanismer er godt beskrevet, men det er alltid en risiko for at slike metoder ikke fanger opp nye endringer i arvestoffet. Andre mer resurskrevende molekylære analyser som inkluderer sekvensering av arvestoffet sammen med resistenstester eller analyser av EC50-verdier er brukt til å oppdage nye mutasjoner som kan gi resistens.
Hvordan tåler soppen fungicider?
Resistensmekanismer er ulike strategier sopp har for å unngå å bli drept av virksomme stoffer i fungicider. Det finnes mange resistensmekanismer, men en av de viktigste er endring i målprotein hos soppen. Et målprotein er et protein hos soppen som det virksomme stoffet i fungicidet binder seg til for så å hindre soppen i å utføre en livsviktig prosess slik at soppen dør eller blir hemmet. Endring i arvestoffet hos soppen gjør at strukturen til målproteinet blir endret slik at det virksomme stoffet i fungicidet ikke klarer å binde seg til målproteinet for å få effekt.
Via en annen mekanismekan sopp unngå at fungicider oppnår ønsket effekt ved å pumpe fungiciders virksomme stoffer ut av cellene før de forårsaker skade. Alle levende organismer har proteinstrukturer i cellemembraner som heter efflukspumper og har som funksjon å transportere giftige stoffer ut av cellene.
Hos noen sopp kan det oppstå endringer i arvestoffet som gjør efflukspumper bedre i stand til å raskt få virksomme stoffer i fungicider ut av cellene, og slik transport kan inkludere virksomme stoffer med ulike biokjemiske virkemåter. Denne typen resistens som forårsakes av efflukspumper gir ofte moderat resistens og kalles for multidrug-resistens (MDR). Det er ikke det samme som multiresistens.
Kryss-resistens og multiresistens
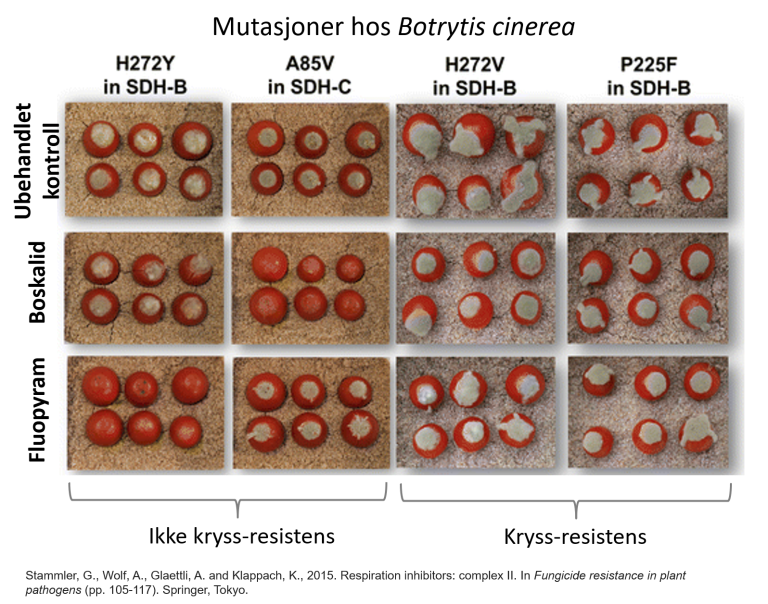
Fungicider tilhører ulike grupper basert på biokjemisk virkemåte, eller hvordan de påvirker livssyklus til soppen. Noen ganger er det kryss-resistens mellom de virksomme stoffene i fungicider som tilhører samme gruppe. Kryss-resistens betyr at resistens utviklet mot et aktivt stoff gir samtidig resistens mot et eller flere andre aktive stoffer i samme kjemiske gruppe. Hos Botrytis cinerea (gråskimmelsopp), for eksempel, er det mutasjoner som gir kryss-resistens mellom boskalid og fluopyram (begge SDHI-fungicider) og mutasjoner som ikke gir kryss-resistens mellom de samme fungicidene.
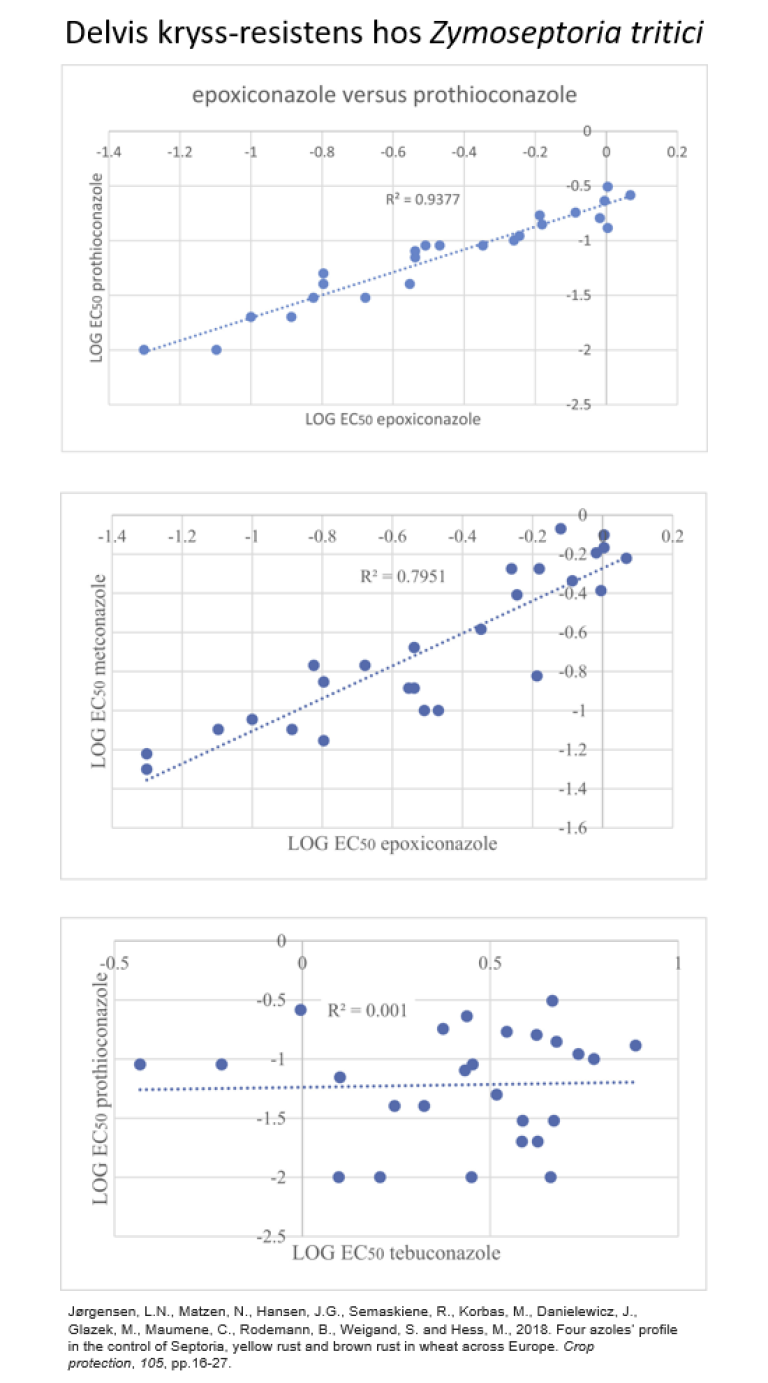
Delvis kryss-resistens kan forekomme mellom fungicider med samme biokjemisk virkemåte også. Figuren under viser korrelasjon mellom gjennomsnitts-EC50-verdier for fire DMI-fungicider hos hvetebladprikksopppen (Zymoseptoria tritici).
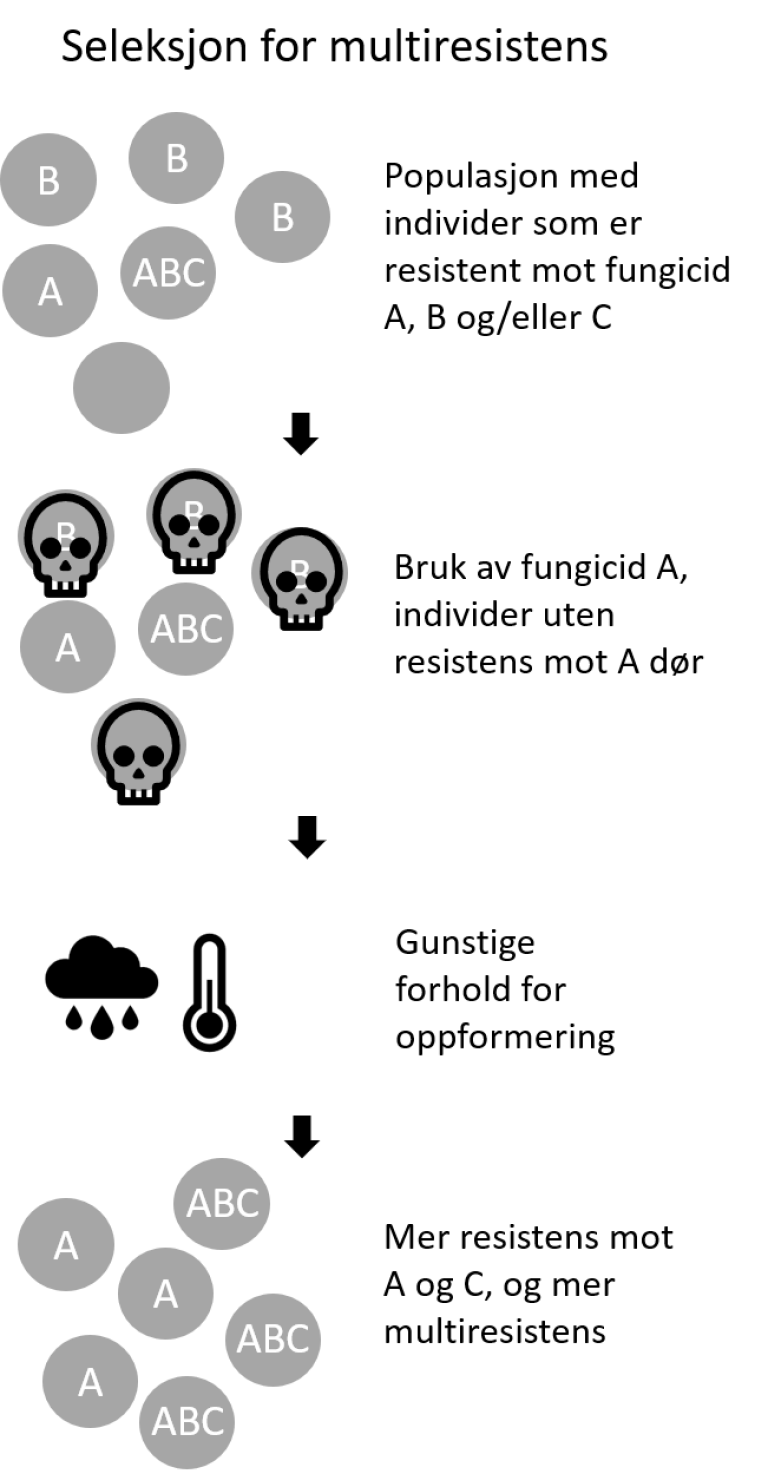
Når enkelte individer blir resistente mot aktive stoffer fra fungicider som tilhører grupper med ulike biokjemiske virkemåter, heter det multiresistens. Når individer i en populasjon er multiresistente, kan det forårsakes av ulike resistensmekanismer og inkluderer ofte mutasjoner i ulike målproteiner som gir en høy grad av resistens mot virksomme stoffer med flere ulike biokjemiske virkemåter. Bruk av et fungicid kan bidra til å opprettholde eller selektere for resistens mot andre fungicider når det er multiresistens.
Hva påvirker risiko for utvikling av fungicidresistens?
Egenskaper hos sopp og fungicider samt agronomiske faktorer påvirker risiko for utvikling av fungicidresistens. (Se lenken til FRAC sin «Pathogen Risk List» for mer informasjon om fungicider og spesifikke patogener.)
Soppens biologi:
Patogene sopparter som har høy genetisk variasjon i populasjonen, har større sjanser for at det tilfeldigvis eksisterer mutasjoner som gir resistens. Slike sopparter vil derfor ha høyere risiko for utvikling av resistens sammenlignet med sopparter som har mindre variasjon. Hver gang sopp formerer seg er det en sjanse for at mutasjoner i arvestoffet hos avkom kan oppstå. Plantepatogene sopparter som formerer seg flere ganger i løpet av en sesong vil derfor ha større sjanser for at det oppstår mutasjoner som gir resistens sammenlignet med sopparter som bare formere seg én gang i løpet av sesongen. Mengde sporer som produseres når soppen oppformerer seg påvirker også resistensrisiko. Arter som har høy produksjon av sporer, har flere sjanser for at det oppstår mutasjoner som gir resistens.
Fungicids virkemåte:
Fungicider som har virksomme stoffer med flere virkesteder gir lavere risiko for utvikling av resistens enn fungicider som har spesifikt virkested. Fungicider med flere virkesteder, eller «multisite»-fungicider på engelsk, påvirker flere steg i livssyklus til soppen. Det er mindre sannsynlig at det forekommer endringer hos soppen som beskytter den mot alle disse virkninger samtidig, uten at disse endringer medfører alvorlig konsekvenser for soppens overlevelse. Ulemper med fungicider med flere virkesteder er at de kan ofte være mer skadelig for andre organismer og mennesker. På grunn av disse ulempene, er det få fungicider med flere virkesteder som er godkjent for bruk i Norge.
Fungicider med virksomme stoffer som har spesifikt virkested, eller «single-site»-fungicider på engelsk, påvirker ett steg i livssyklusen til soppen. En enkelt mutasjon hos soppen kan være tilstrekkelig for å endre målproteinet nok til at det virksomme stoffet i fungicidet mister effektiviteten. Selv om fungicider med spesifikt virkested er mer resistensutsatt, så er de også mer skånsom overfor andre organismer og mennesker nettopp på grunn av at de har spesifikk virkning mot sopp. De fleste fungicider som er godkjent for bruk i Norge inneholder virksomme stoffer med spesifikt virkested.
Agronomiske faktorer:
Agronomiske faktorer inkluderer alle aspekter av produksjon som påvirker smittepress lokalt. Faktorer kan inkludere smittekilder, værforhold, gjødsling, vanning, kulturtiltak, og sortsvalg. Alt som gjør plantebestanden mer gunstig for overlevelse og oppformering av sopp, bidrar til økt risiko for resistensutvikling.
Hvordan unngå/utsette oppbygging av fungicidresistens?
Hver behandling med fungicider øker seleksjonspresset for fungicidresistens. Å bruke integrert plantevern og begrense bruken av fungicider til når det virkelig trengs er viktig for å unngå eller utsette oppbygging av fungicidresistens. Når fungicider må brukes er det viktig å unngå ensidig bruk av fungicider med samme biokjemiske virkemåten. Det er ikke alltid tilstrekkelig å veksle mellom produkter ettersom noen produkter inneholder ulike aktive stoffer som tilhører samme kjemisk gruppe og det kan være kryss-resistens. Unngå bruk av midler der det er påvist resistens på et risikonivå hos soppen. Oppsøk råd om antiresistensstrategier som er spesifikke for ulike kulturer eller produksjonssystemer.
Tjenester
Plantevernleksikonet
Nettside med nyttig og relevant informasjon om biologi og bekjempelse av planteskadegjørere (ugras, sykdommer og skadedyr). Du finner også informasjon om biologien til en del nyttedyr av plantevernbetydning.
Plantevernguiden er en nettbasert tjeneste som gir deg en samlet oversikt over godkjente kjemiske og biologiske plantevernmidler. Tjenesten er utviklet i et samarbeid mellom Mattilsynet og NIBIO.
Det er per i dag påvist resistens eller nedsatt følsomhet mot kjemiske plantevernmidler hos flere skadedyr, plantepatogener og ugras i norske jord- og hagebrukskulturer. Hos skadedyr er resistens mot pyretroider og nedsatt følsomhet for tiakloprid vanlig hos rapsglansbille i oljevekster. Resistens mot pyretroider er påvist hos ferskenbladlus og potetsikade fra potet, gulrotsuger fra gulrot, ferskenbladlus fra persille, kålmøll og ferskenbladlus fra kålvekster, jordbærsnutebille fra jordbær, og ferskenbladlus, bomullsmellus, veksthusmellus og sør-amerikansk minerflue fra veksthus. Det er også funnet resistens mot pirimikarb hos ferskenbladlus og nedsatt følsomhet for imidakloprid hos ferskenbladlus og bomullsmellus. I jordbær og bringebær er det indikasjoner på begynnende resistensutvikling mot flere av middmidlene. Hos plantepatogener er resistens mot QoI-fungicider
påvist hos gråskimmel fra jordbær, bringebær og gran i skogplanteskoler, hos mjøldoggsopper i jordbær og veksthusagurk, og hos bladflekksopper i hvete. Resistens mot triazoler er funnet i flere bladflekksopper i hvete. Resistens mot hydroksyanilid- og SDHI-er utbredt hos gråskimmel fra jordbær og bringebær, og i skogplanteskoler er det påvist resistens mot tiofanater.....