Thiago Inagaki
Forsker
Biografi
Bachelor- og mastergrad i agronomi og doktorgrad i naturvitenskap (Dr. rer. nat.) med fokus på jordvitenskap. Forskningen fokuserte på effekten av arealbruk og forvaltning på jordas C-innhold, og mekanismer for beskyttelse av organisk materiale i jorda gjennom organo-mineralforbindelser. Ledet prosjekter som krysser jordkvalitet, bærekraft og geovitenskap ved hjelp av avanserte metoder, inkludert 13C-NMR, SEM/TEM, STXM, NanoSIMS, FIB-SEM og stabile isotoper (13C og 15N).
Forfattere
Raphael Silva Thiago Inagaki Thomas Trentin Laura Péres Cléber Jesus Luis Alleoni Matheus SoaresSammendrag
Highly reactive, fine-particle biochars are particularly effective in recarbonizing degraded soils, but they can be more vulnerable to fire degradation due to higher surface area. How biochar particle size influences its persistence in the environment remains poorly understood, particularly in regions affected by fire events such as the Cerrado. In this study, we investigated the stability of micrometric (200 µm) and nanometric (<50 nm) açaí seed biochar incubated in an Oxisol for 180 days, followed by simulated burning using a propane–butane torch. Biochar amended soil had higher total C and nitrogen (N) contents, with a 43% increase for microbiochar and 88% for nanobiochar in the mineral-associated organic matter (MAOM) fraction, indicating incorporation into stable pools. Burning reduced C associated with the particulate organic matter (POM) more when amended with microbiochar than with nanobiochar, demonstrating their vulnerability fire. An increase, up to fivefold, in dissolved organic carbon (DOC) and nitrogen (DN) contents occurred after burning. XRD data showed no mineralogical differences among samples, while FTIR indicated loss of polysaccharides after burning. TGA revealed that microbiochar was more thermally stable under unburned conditions but lost stability after burning. In contrast, nanobiochar was less stable in unburned soil but became more stable following burning, resulting in lower mass loss. The reduced mass loss in the burned control is attributed to its ~40% lower soil C content. Biochar’s particle size governs carbon stability, fire vulnerability, and post-burn soil resilience, thereby using nanobiochar for fire-prone areas act as strategy improving the management of agricultural soils.
Forfattere
Thiago InagakiSammendrag
Det er ikke registrert sammendrag
Forfattere
Thiago InagakiSammendrag
Det er ikke registrert sammendrag

Divisjon for miljø og naturressurser
Conservation of Biodiversity in China in the light of Climate Change
Climate change is becoming an increasingly important pressure on biodiversity, which adds to the burden of other drivers of loss of biodiversity causing negative effects on ecosystems and species

Divisjon for miljø og naturressurser
AgriCascade
Cascading recycling of organic N-sources with next-generation biochar fertilizer for Norwegian agriculture

Divisjon for miljø og naturressurser
AgriCascade
AgriCascade: Gjenvinning av organisk nitrogen med forbedret biokull for gjødslings- og miljøgevinster i norsk landbruk
Divisjon for miljø og naturressurser
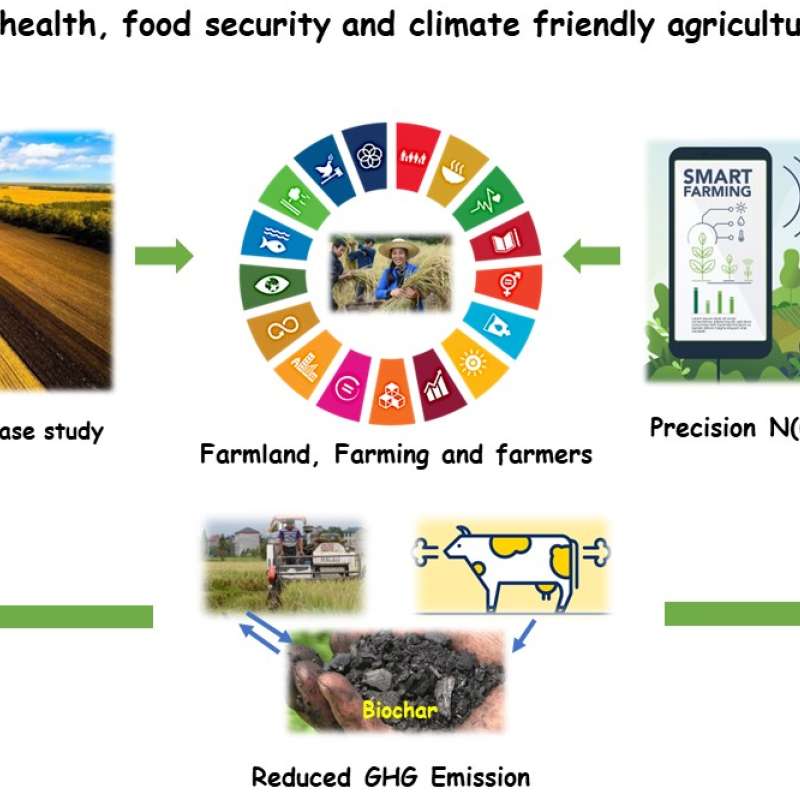
Sinograin III: Smart agricultural technology and waste-made biochar for food security, reduction of greenhouse gas (GHG) emission, and bio-and circular economy
The Sinograin III project’s overall objective is to contribute to the UN SDGs by widely implementing precision agriculture technologies and application of “waste-to-value” biochar products to achieve sustainable food production with minimized GHG emission, improve soil fertility and promote green growth/zero waste in modern agriculture in China.
Divisjon for miljø og naturressurser
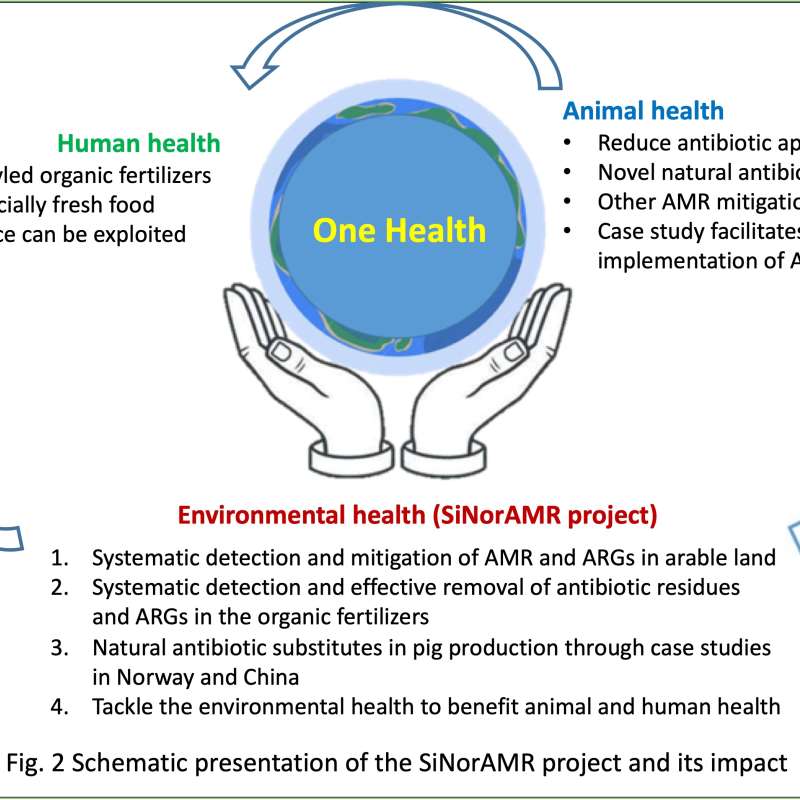
SiNorAMR
Full title: Collaborative and Knowledge-building Project Collaborative Project Systematic detection and mitigation of antimicrobial resistance in soil environment and animal health contributing to human health (SiNorAMR)